Introduction
As an introduction, watch this movie.
If the movie does not play in this window, or you would like to see it in a window of alternate size, download it from this link.
Release of Neurotransmitter from a Nerve Terminal
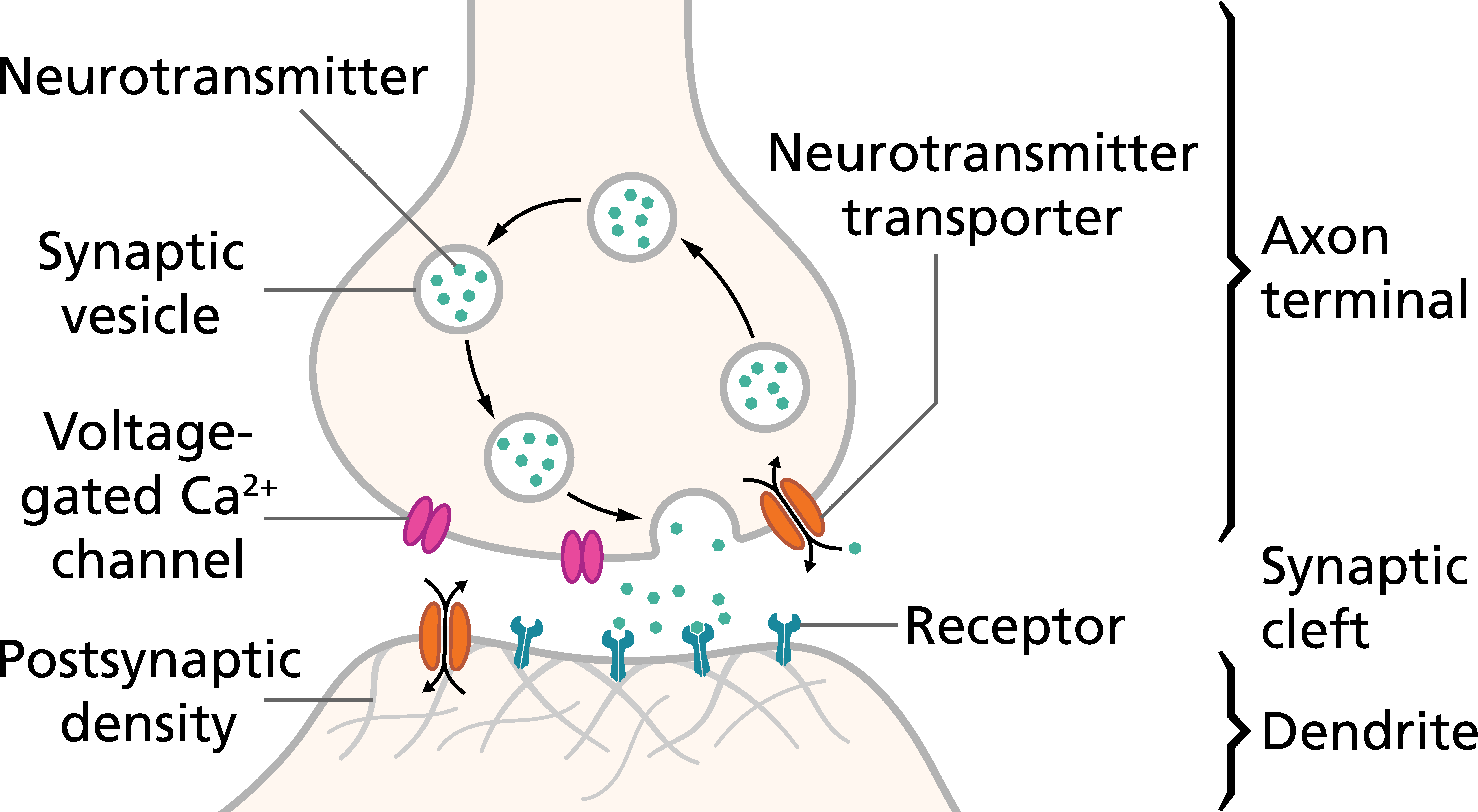
Neurotransmitters are chemicals that transmit signals from one neuron to the next. Neurotransmitters are packaged into vesicles located in the nerve terminal. When an action potential depolarizes the nerve terminal, voltage-gated Ca2+ channels are opened, allowing Ca2+ to enter the terminal. When the calcium ions enter the presynaptic terminal, they bind with special protein molecules on the inside surface of the presynaptic membrane, called release sites. This binding in turn causes the release sites to open, allowing a few transmitter vesicles to release their transmitter into the synaptic cleft (space between a nerve terminal and the dendrites or soma of another neuron). The released neurotransmitter binds to receptors on the dendrite or soma of the postsynaptic neuron.
Let's make sure the terminology is clear:
- Synapse: a junction between two nerve cells, where information is transmitted chemically across a small gap (or synaptic cleft)
- Presynaptic neuron: a neuron that releases a neurotransmitter
- Postsynaptic neuron: a neuron that has receptors for the neurotransmitter released from a presynaptic neuron
Postsynaptic Actions of Neurotransmitters
When a neurotransmitter binds to a receptor on a postsynaptic neuron, one of two things usually happens:
- A ligand-gated ion channel opens, resulting in the passage of ions across the membrane. This is called an ionotropic response. Such responses have a rapid onset and a rapid termination. These receptor-channel complexes are called ionotropic receptors.
- A G-protein-mediated second messenger system is activated, resulting in biochemical modifications that can alter ion channel permeability and many other aspects of cellular function. This is called a metabotropic response. Such responses have a slow onset and a slow termination. These receptors are called metabotropic receptors.
Excitatory and Inhibitory Neurotransmitters
Although there are many distinct neurotransmitters, as will be described in subsequent modules, they can be divided into two general groups: excitatory and inhibitory.
- Excitatory neurotransmitters make the postsynaptic neuron more likely to fire. This occurs by depolarizing the postsynaptic neuronal membrane.
- Inhibitory neurotransmitters make the postsynaptic neuron less likely to fire. This usually occurs through hyperpolarization or stabilization of the postsynaptic neuronal membrane.
Postsynaptic Potentials
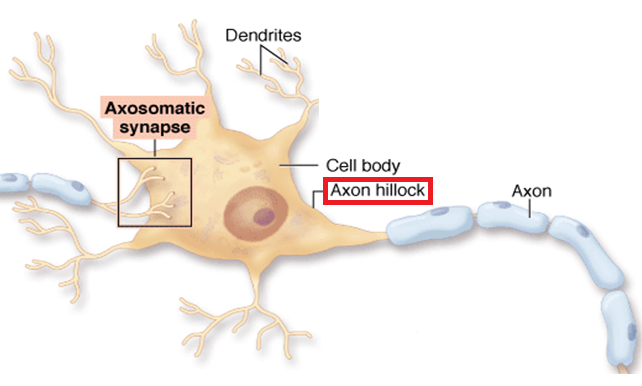
The axon hillock is constantly "summating" the thousands of EPSPs and IPSPs that are generated in the dendrites and soma. If this summation amounts to a depolarization of sufficient magnitude, the activation gates of the voltage-gated Na+ channels will open, an action potential will occur, and the neuron will release neurotransmitter onto the neurons it synapses upon. When the depolarization at the axon hillock is smaller than the threshold required to open the activation gates of the voltage-gated Na+ channels, the neuron will not communicate with other neurons down the line. Whether or not a neuron communicates with other neurons all comes down to the membrane potential at the axon hillock at a particular time.
Some Synapses Are Different
Most synapses are axo-dendritic (i.e., the synapse is on a dendrite) or axo-somatic (i.e., the synapse is on the neuronal soma), as described above. However, other synaptic configurations also exist. One such configuration is the axo-axonic synapse, where an axon of one neuron synapses on the axon of another neuron.
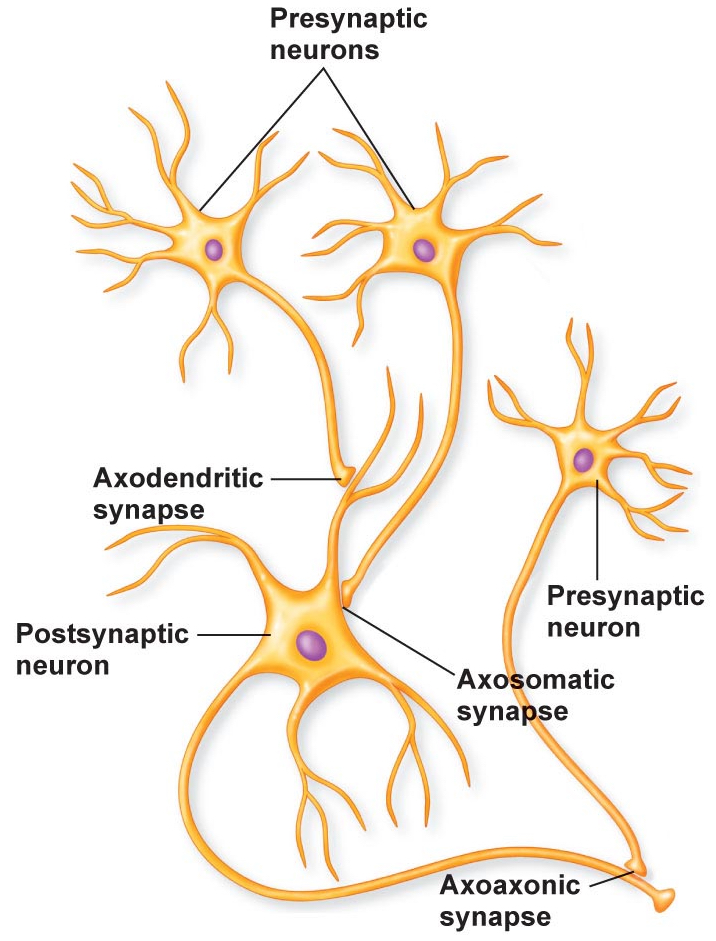
What is the function of an axoaxonic synapse? One common role of such synapses is producing presynaptic inhibition. The axoaxonic synapse can activate a second messenger system that affects the voltage-gated Ca2+ channels at the nerve terminal, so they allow less Ca2+ to enter the nerve terminal after an action potential occurs. As a result, the neuron releases less neurotransmitter following an action potential. Axoaxonic synapses thus provide a mechanism to regulate how much a neuron communicates to other neurons. Afferent fibers entering the spinal cord are subject to presynaptic inhibition via axoaxonic synapses.
A similar phenomenon is presynaptic facilitation. Second messenger systems triggered through axoaxonic synapses can cause more calcium to enter the nerve terminal when an action potential occurs, so a neuron releases more neurotransmitter. It is believed that presynaptic facilitation plays a key role in learning and memory.
Although we normally think of neurotransmitters binding to postsynaptic receptors, some nerve terminals have autoreceptors for the neurotransmitter they release. Often these autoreceptors play a homeostatic role, and serve to adjust the amount of neurotransmitter released subsequently. In other words, autoreceptors serve to assure that a nerve terminal is not releasing too much, or too little, neurotransmitter.
Let's review these concepts by watching a movie from the KhanAcademy.
If the movie does not play in this window, or you would like to see it in a window of alternate size, download it from this link.
Categories of Neurotransmitters
The most common neurotransmitters are amino acids, particularly glutamate, GABA (gamma-aminobutyric acid), and glycine. The actions of these amino acids will be discussed in subsequent modules. Glutamate is an excitatory neurotransmitter, and is released by most sensory afferents as well as neurons in the central nervous system. GABA and glycine are inhibitory neurotransmitters; glycine is mainly released from spinal cord neurons, whereas GABA is released from neurons in many brain areas. In fact, GABA is the most common inhibitory neurotransmitter present in the central nervous system.
Another very common neurotransmitter is acetylcholine. Acetylcholine is released by neurons in many brain regions, as well as by motoneurons, sympathetic and parasympathetic preganglionic neurons, and parasympathetic postganglionic neurons. Acetylcholine is mainly an excitatory neurotransmitter, although there are some examples where it has an inhibitory role (e.g., parasympathetic postganglionic nerve fibers that lower heart rate).
Another group of important neurotransmitters are the monoamines, chemicals that contain one amino group that is connected to an aromatic ring by a two-carbon chain (-CH2-CH2-). The monoamine neurotransmitters include dopamine, norepinephrine, and serotonin. Monoamine neurotransmitters typically bind to G-protein coupled receptors, such that they can produce a wide variety of effects in the postsynaptic neuron. Many neurons that release monoamine neurotransmitters branch extensively, such that each neuron makes synapses on a large number of postsynaptic neurons. Many neurological and psychiatric diseases are related to the monoamine neurotransmitters.
Nitric oxide (NO) is released by nerve terminals in areas of the brain responsible for long-term behavior and memory. NO is different from other small-molecule transmitters, as it is very lipophilic and cannot be stored in the nerve terminal. Instead, it is synthesized as needed and then diffuses out of the presynaptic terminals to affect adjacent neurons. NO usually does not alter the membrane potential, but instead changes intracellular metabolic functions that modify neuronal excitability for seconds, minutes, or perhaps even longer. NO is also an important signaling molecule in the cardiovascular system, as you learned in the Cardiology Course.
A variety of peptides called neuropeptides can also act as neurotransmitters, but they usually have very specialized functions. For example, some hypothalamic neurons release peptides that control the release of hormones from the anterior pituitary. In some cases, a peptide hormone is co-released with one of the "classical" neurotransmitters discussed above.
Let's review these concepts by watching a movie from the KhanAcademy.
If the movie does not play in this window, or you would like to see it in a window of alternate size, download it from this link.